Порфириновое кольцо представляет собой плоскую рамку
Гемы
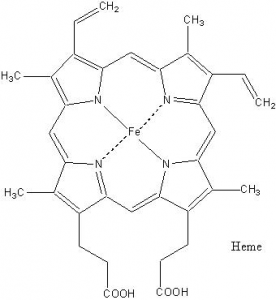
Ключевыми элементами элементами комплекса IV, играющими основную роль в переносе электронов, являются особые простетические группы – гемы.
{kind=link}
Гем представляет собой плоскую структуру, базой которой является порфириновое кольцо, включающее, в частности, четыре расположенные в вершинах квадрата атома азота, каждый из которых опирается на два атома углерода. В центре гема находится катион железа, связанный ковалентными связями с двумя диагонально расположенными атомами азота, и имеющий возможность отдавать и получать электрон, переходя из состояния Fe2+ в Fe3+ и обратно.
Колебания атомов в геме
Атомы всех молекул клетки совершают колебания в различных направлениях с амплитудой, определяемой температурой и наличием связей с другими атомами.
В геме, как и во всякой другой плоской структуре, наибольшую амплитуду имеют колебания атомов в направлении нормали к плоскости.
Сила, возвращающая атом в равновесное положение, определяется величиной смещения из равновесного положения (в частности, в случае упругой связи, пропорциональна величине смещения). Если какой-либо атом сместился из равновесного положения в направлении нормали к плоскости на малую величину Δ, то расстояния от этого атома до других атомов, с которыми он связан, меняется на величину второго порядка малости по отношению к Δ или, что то же самое, смещение от положения равновесия имеет второй порядок малости. Это означает, что при наличии упругой связи возвращающая сила имеет также второй порядок малости.
Наибольшую из всех атомов амплитуду колебаний в направлении нормали к плоскости имеет катион, расположенный в центре структуры, а амплитуды колебаний других атомов убывают по мере удаления от центра к краям гема.
Собственные частоты малых колебаний системы атомов гема зависят от способа закрепления краёв гема и приблизительно одинаковы для всех гемов.
Предполагаемый механизм передачи энергии в хемо-электрических преобразователях
В комплексе IV имеются два гема, один из которых гемов находится в цитохроме а3 на активном центре, осуществляющем экзергическую реакцию, а второй – в цитохроме а на положительной стороне мембраны. Будем считать катион железа, принадлежащий цитохрому а3 первым, а цитохрому а – вторым.
Предлагаемая модель основана на двух предположениях:
- плоскости обоих гемов приблизительно параллельны поверхности мембраны;
- центры гемов находятся приблизительно один под другим.
В этом случае оба катиона железа совершают колебания, главным образом, в направлении нормали к поверхности мембраны с одной и той же частотой. Сдвиг по фазе межуду колебаниями двух гемов, как будет видно далее, должен быть близок к π.
Под действием внутримембранного макрополя все атомы гема и, в частности, катионы железа поляризуются, так что их электронные облака смещаются к положительной стороне мембраны.
Приближаясь в ходе колебаний ко второму катиону, находящемуся в этот момент в состоянии Fe2+, первый катион создаёт в окрестности второго электрическое микрополе, противоположное по направлению внутримембранному макрополю и смещающему его электронное облако к себе. В результате при достаточном сближении катионов их электронные облака перекрываются, т.е. некоторые электроны часть времени находятся у ядра одного атома и часть времени – у другого. В этих условиях при расхождении ядер катионов один из таких общих электронов остаётся у более сильного катиона. При этом первый катион переходит в состояние Fe2+, а второй – в состояние Fe3+.
Эти колебания поддерживаются толчками, получаемыми после каждого акта экзергической реакции в тот момент, когда первый катион отдал свой электрон и перешёл в состояние Fe3+.
Работа над файлом должна быть продолжена
Энергия электромагнитного излучения
При использовании энергии электромагнитного излучения выделению энергии предшествует поглощение фотона определённой группой атомов пигментной молекулы (более подробно см. Фотосинтез). Электрон, перешедший в результате поглощения фотона на более высокий энергетический уровень, может отдать полученную энергию двумя способоми
//////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
Из статьи Фотосинтез в Википедии
Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и кислорода порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10—12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносов электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции H20 → O2 E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.
Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачу энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10-10-10-12 сек., расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
Солнечное излучение поглощается в состоящих из нескольких сотен пигментных молекул светоприёмных комплексах (СПК), которые в случае простого цикла зарядки являются частью ФС1.
Поглощённые кванты солнечного излучения преобразуются в СПК в локализованные возбуждения групп атомов, способные перемещаться по энергопередающим линиям к фотореакционному центру (ФРЦ) – молекуле хлорофилла, находящейся на пересечении СПК и КПЭ ФС1.
Согласно теории Фёрстера, выводы которой согласуются с более поздними исследованиями А.С. Давыдова, эти возмущения можно рассматривать как экситоны.
Энергия экситона постепенно рассеивается в окружающую среду и может быть передана только на небольшие расстояния.
Структура фотосистем
* Фотосистемы представляют собой пигментно-полипептидные комплексы, встроенные в мембраны тилакоидов.
* Растения и цианобактерии имеют в каждом тилакоиде фотосистемы двух типов: фотосистемы-1 (ФС1) и фотосистемы-2 (ФС2).
* Каждая фотосистема независимо от типа имеет два функциональных комплекса молекул – комплекс подъёма электронов (КПЭ), расположенный по оси фотосистемы, и светопринимающий комплекс (СПК), обрамляющий КПЭ снаружи.
* Как ФС1, так и ФС2 цианобактерий имеют в составе СПК несколько сотен расположенных веерообразно вокруг КПЭ молекул пигментов – хлорофиллов а и b, а также различных фикобилинов (у растений вместо фикобилинов используются каротиноиды).
* Молекулы, входящие в состав КПЭ, выстроены в цепочку, пересекающую тилакоидную мембрану.
* Общей частью КПЭ и СПК является молекула хлорофилла, образующая фотореакционный центр (ФРЦ).
ФРЦ
* ФРЦ фотосистем обоих типов состоит из молекул хлорофилла, содержащих порфириновое кольцо, в центре которого находится атом магния.
* Атом магния имеет в наружной оболочке два электрона, с помощью которых он образует ковалентные связи с двумя атомами азота.
* При потере одного из электронов наружной оболочки у атома магния он теряет связь с одним атомом азота, но остаётся связанным с другим и, значит, не выпадает из молекулы.
* Молекула хлорофилла в электрически нейтральном состоянии может ионизироваться, отдавая один электрон атома магния, и, следовательно, играть роль донора электрона.
* В ионизированном состоянии молекулы хлорофилла её электрическое поле аппроксимируется полем элементарного положительного точечного заряда, расположенного в ядре атома магния.
* Рекомбинируя, ионизированная молекула хлорофилла может играть роль акцептора электрона.
Нижняя часть КПЭ
* Молекулы «нижней» части КПЭ, примыкающей к ФРЦ со стороны люмена, включают атомы, которые могут проявлять положительную валентность больше двух и, следовательно, терять более одного электрона, не теряя связь с молекулой.
* Благодаря атомам с высокой положительной валентностью, молекулы нижней части КПЭ могут, оставаясь катионами, попеременно терять и присоединять один электрон, переключаясь из состояния донора в состояние акцептора.
Верхняя часть КПЭ
* Молекулы верхней части КПЭ в электрически нейтральном состоянии способны играть роль акцептора для электронов, поступающих снизу, превращаясь в анион, а в ионизированном состоянии – отдавать электрон «наверх», играя роль донора.
// ФС2 цианобактерий содержит 17 полипептидных субъединиц, включающих в себя в сумме 45 000 атомов.
Зарядка со сложным циклом
Принципиальная схема и структурная реализация процесса
* В структурной реализации процесса зарядки со сложным циклом определяющую роль играют три встроенных в тилакоидную мембрану макромолекулярных комплекса: фотосистема-2 (ФС2), ЦК и ФС1.
* Сложный цикл включает два сопряженных цикла.
Цикл 1
1) Водорасщепляющий комплекс —примыкающая к люмену часть ФС2 – захватывает из люмена молекулу воды и разлагает её на атом кислорода О, два протона Н+ и два электрона е– .
// Разложение молекулы воды требует затрат энергии ЕWS.
2) Атом О и два протона Н+ поступают в пул люмена, а два электрона с помощью принадлежащего ФС2 КПЭ переносятся из люмена в строму.
// Положительный заряд люмена увеличился на две элементарных единицы.
3) На обращённой в строму стороне ФС2 перенесенные в строму электроны принимаются молекулой пластохинона PQ, превращающейся при этом в дважды заряженный анион PQ2–.
4) С помощью анионов PQ2–, являющихся, как принято считать, подвижными молекулами, перемещающимися в ходе пассивной диффузии, два электрона переносятся к стромальной стороне ЦК на n-сайт.
5) С участием ЦК анион PQ2– вступает в реакцию с извлечённым из пула стромы гидронием НзО+, что приводит к появлению молекул воды Н2О и пластохинола PQН2.
// Превращение двух противоположно заряженных частиц в две нейтральные частицы связано с выделением энергии.
// Отрицательный заряд стромы увеличился на две элементарных единицы.
6) Принято считать, что молекула пластохинола подвижна и в процессе пассивной диффузии переносится внутри тилакоидной мембраны со стромальной стороны на люменальную, где закрепляется на р-сайте.
7) На люменальной стороне ЦК входящие в него ферменты осуществляют 4-х стадийный процесс отбора двух атомов водорода от молекулы РQН2 и расщепления их на протон и электрон.
// Два свободных протона переходят в пул люмена, а электроны остаются связанными на белковых молекулах ЦК.
// Суммарный заряд люмена при этом не изменяется, т.к. оставшиеся электроны находятся в той же зоне.
8) Принято считать, что молекула пластохинона, получившаяся после отбора от от пластохинола двух атомов водорода, переходит в процессе пассивной диффузии внутри тилакоидной мембраны от люменальной стороны ЦК к стромальной стороне ФС2.
Цикл 2
Пошаговое распределение затрат энергии
* Перемещение электронов из люмена в строму является многошаговым процессом, на каждом шаге которого один электрон «поднимается» от «нижележащей» молекулы КПЭ к «вышележащей».
* Энергия Еliftk, затрачиваемая на k-ой ступени подъёма для преодоления сил электрического макрополя, пропорциональна высоте ступени Δhk = zk+1 – zk для рассматриваемого шага:
Еliftk = Δhk/h ∙Еlift. (2)
* Одна часть пути, который проходит электрон при подъёме от люмена к строме, лежит ниже ФРЦ, а другая часть – выше.
ФРЦ – точка бифуркации потока энергии в КПЭ
* Поступивший от ССК к молекуле ФРЦ солитон передаёт ей часть своей энергии Еsol0, которая, по-видимому, возбуждает атом магния в центре порфиринового кольца.
* Один из электронов валентной оболочки атома магния переходит на столь высокий уровень, что он может быть перехвачен атомом вышележащей нейтральной молекулой А, способной играть роль акцептора.
* Молекула ФРЦ теряет энергию Еdet, затраченную на ионизацию атома магния, и превращается в катион.
* Присоединивший электрон акцептор А превращается в анион Ан.
* Чтобы обеспечить подъём электрона на всём пути от люмена к строме, солитоны, приходящие от СПК к ФРЦ должны разделяться на две части, одна из которых направляется в верхнюю часть КПЭ и включается в восходящий поток энергии, а вторая – в нижнюю часть и включается в нисходящий поток.
Энергетические затраты в восходящем потоке
* Восходящий поток энергии в верхней части КПЭ обеспечивает последовательный подъём электронов от нижележащего аниона Ан к вышележащему нейтральному акцептору А. При этом на k-ом шаге подъёма:
- Ан возбуждается, получив энергию Еsolk от пришедшего к нему солитона;
- один из электронов Ан переходит на столь высокий уровень, что может быть перехвачен положительным полюсом одного из членов мультипольного разложения поля А;
- Ан теряет энергию Еdetk, унесенную утраченным электроном (главным образом, работа отрыва электрона от положительного полюса мультипольного разложения его микрополя), и превращается в нейтральную молекулу, которая может играть роль акцептора в случае прихода нового возбуждения;
- А, получив электрон, превращается в анион, а его энергия увеличивается на величину работы Еdetk+1, произведенной его микрополем при присоединении электрона, и уменьшается на величину работы Еliftk, затраченной на подъём электрона в макрополе;
* На последнем шаге восходящего потока энергии превращается в анион находящийся в строме конечный акцептор.
* Отбор электрона от аниона конечного акцептора в отрицательно заряженную строму невозможен без использования специального механизма.
Энергетические затраты в нисходящем потоке
* Нисходящий поток энергии обеспечивает последовательный подъём электрона от нижележащего атома донора Д к вышележащему катиону Кат. При этом на k-ом шаге подъёма:
- атом Д возбуждается, получив энергию Еsolk от пришедшего к нему солитона;
- валентный электрон атома Д переходит на столь высокий уровень, что может быть перехвачен микрополем Кат;
- атом Д теряет энергию Еdetk, затраченную на ионизацию, и может играть роль катиона для следующего шага;
- лежащий выше Д катион Кат, притягивая к себе отданный Д электрон, рекомбинирует и может играть роль донора для следующего цикла;
- при рекомбинации Кат вновь получает энергию Еdetk+1, потерянную ранее при ионизации, и теряет энергию Еliftk, затраченную на подъём электрона в макрополе;
* На последнем шаге нисходящего потока энергии отбирается электрон от находящегося в люмене первичного донора.
* Получить электрон для рекомбинации первичного донора из положительно заряженной стромы невозможно без использования специального механизма.
Механизм регенерации первичных доноров и конечных акцепторов
Водорасщепляющий комплекс (ВРК) – устройство для регенерации первичного донора ФС2
* Роль первичного донора для ФС2 играют два нижние атома Mn четырёхатомного марганцевого кластера.
* Входящие в марганцевый кластер атомы постоянно ионизированы и, последовательно получая и теряя электрон, меняют своё состояние от Mn+ до Mn2+.
* Источником электронов для Mn2+ является молекула воды.
* Получение электронов от воды обеспечивает примыкающий к ФС2 со стороны люмена белковый водорасщепляющий комплекс (ВРК).
* ВРК захватывает из пула люмена молекулу Н2О и расщепляет её на один атом кислорода О, два протона Н+ и два электрона е–.
* К моменту расщепления нижние атомы марганца находятся в состоянии Mn2+, что позволяет им одновременно «затолкнуть» протоны Н+ в отталкивающий их люмен и присоединить к себе электроны е–, совершив работу по их поднятию против сил макрополя.
* ВРК затрачивает энергию, получаемую от пришедшего солитона, на расщепление молекулы воды.
* Особенности структуры КПЭ2 обеспечивают одновременный подъём обоих выделившихся в ВРК электронов.
* Оставшийся нейтральный атом свободного кислорода без сопротивления переходит в пул люмена, после чего ВРК размещает на освободившемся посадочном месте новую молекулу воды.
Энергия Е2, затрачиваемая в ФС2 при увеличении заряда конденсатора на одну элементарную единицу, включает в себя, кроме энергии Еlift, необходимой для подъёма электрона с положительной обкладки на отрицательную, также половину энергии ЕWS, необходимой для расщепления молекулы воды:
Е2 = Еlift + ½ ЕWS. (5)
Все биологические макромолекулы (полисахариды, белки и нуклеиновые кислоты), являются полимерами, состоящими их одномерных цепочек более простых блоков – мономеров.
В частности, входящие в состав светособирающих комплексов пигментные молекулы имеют длинный хвост, состоящий из одинаковых углеводородных мономеров.
Как было показано в работах А.С. Давыдова и др. авторов, выполненных в последней четверти ХХ века, в биополимерах возможно распространение переносящих энергию нелинейных локализованных возбуждений – солитонов, обладающих определённой пространственной конфигурацией и скоростью перемещения.
- Вполне вероятно, что распространение солитонов по полимерным цепям является главным способом переноса энергии в клетках живых организмов.
Если сделанные предположения верны, то биологические макромолекулы состоят из активной части (в случае хлорофилла – порфиринового кольца), в которой происходит преобразование энергии электромагнитного излучения или окислительно-восстановительного потенциала молекул в локализованное возбуждение – солитон, и полимерной линии передачи энергии в виде солитонов.
Дата последнего обновления: 2008-04-28