Мембраны как диэлектрические прокладки клеточных конденсаторов
Все биологические мембраны обладают хорошими электроизолирующими свойствами благодаря двойному липидному слою,
С обеих сторон каждой такой мембраны находятся электролиты (т.н. проводники второго рода), которые могут быть заряжены отрицательно или, на языке химии, иметь щелочную реакцию (рН > 7) или положительно иметь кислую реакцию, т.е. рН < 7.
В каждой клетке имеются специальные энергетические мембраны, в которых вмонтировано оборудование для переноса заряда с одной стороны мембраны на другую и создания тем самым в соответствующих электролитах зарядов противоположных знаков.
С точке зрения электротехники совокупность энергетической мембраны и противоположно заряженных электролитов по её сторонам представляет собой конденсатор, в котором роль обкладок играют электролиты.
Таблица. Основные виды энергетических мембран и клеточных конденсаторов
| Мембрана | Отрицательная обкладка | Положительная обкладка |
| Клеточная мембрана | Цитоплазма | Слой внешней среды, непосредственно примыкающий к клетке |
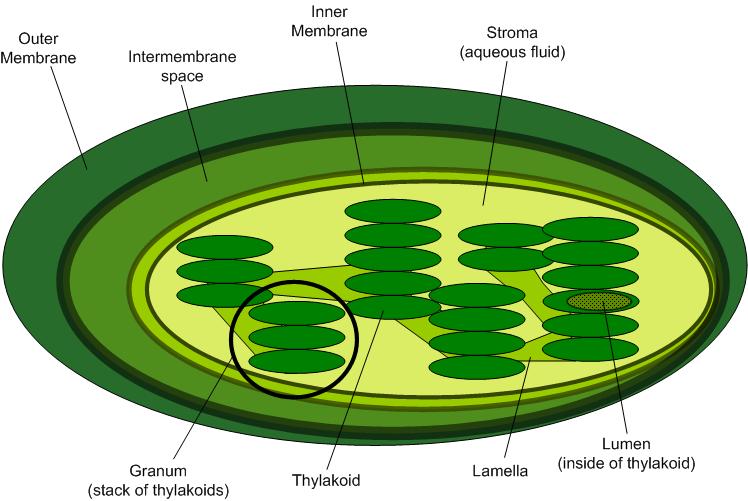
| Тилакоидная мембрана | Строма | Люмен |
| Внутренняя мембрана митохондрий | Матрикс | Внутримембранное пространство |
Схематическое изображение клеточной мембраны из Википедии.
{kind=link}
Структура хлоропласта (Википедия)
{kind=link}
Структура митохондрии (Википедия)
{kind=link}
Схематическое изображение тилакоидов (зелёные) в цианобактерии Synechocystis
{kind=link}
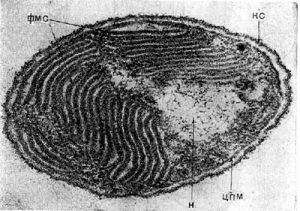
Ультратонкие срезы клеток Ectothiorhodospiга shaposhnikovii.
КС — клеточная стенка, ЦПМ — цитоплазматическая мембрана, Н — нуклеоид, ФМС — фотосинтезирующие мембранные структуры.
Увел. X 40 000. (Фото из статьи «Фототрофные бактерии»).
Внутримембранное электрическое поле и запасённая энергия
Обозначения:
А – площадь поверхности мембраны, м2;
Е – напряжённость электрического поля, В/м;
V – разность потенциалов на обкладках, В;
Δ – толщина мембраны, м; все биологические мембраны имеют примерно одинаковую толщину ≈ 7 нм.
ε – диэлектрическая проницаемость мембраны, Ф/м; для приближённых оценок можно принять ε = ε0, где ε0 = 8,85 ∙ 10–12 Ф/м – электрическая постоянная.
Ω – запасаемая в конденсаторе энергия электрического поля, Дж;
ω – объёмная плотность энергии электрического поля, Дж/м3.
Практически всё поле биологического конденсатора сосредоточено в мембране и имеет напряжённость
Е = V /Δ. (1)
С учётом (1) для объёмной плотности энергии электрического поля получается формула
ω = ½ ε Е2 = ½ ε V 2/Δ 2. (2)
Умножив обе части (2) на объём мембраны АΔ, получаем выражение для запасаемой в конденсаторе энергии электрического поля
Ω = ω АΔ = ½ ε АV 2/Δ = α АV 2, (3)
где α ≈ 1.3 10–3 Ф/м2.
Максимально возможная величина V* разности потенциалов определяется электрической прочностью мембраны, также примерно одинаковой для всех биологических мембран. Таким образом, максимально возможная запасённая энергия Ω* пропорциональна площади поверхности мембраны А.
Способы создания заряда на обкладках и соответствующие затраты энергии
Любой водный раствор, кроме нейтральных молекул Н2О и ионов растворённых веществ, содержит также положительные ионы гидрония НзО+ (молекула воды с присоединённым протоном) и отрицательные гидроксильные ионы ОН– (молекула воды с отщепленным протоном).
Свободные протоны в воде могут существовать лишь ничтожно малое время, т.к. легко соединяются с молекулами воды, образуя гидроний.
В электрически нейтральном растворе число катионов и анионов одинаково, т.е., в частности, в чистой воде одинаково число ионов гидрония и гидроксила. Водный раствор можно зарядить положительно или отрицательно, создав преобладание соответственно ионов гидрония или гидроксила, т.е., в конечном счёте, создав избыток или недостаток протонов по отношению к электрически нейтральному состоянию. Это означает, что увеличить заряд обкладок конденсатора можно следующими тремя способами.
- Выделение протона на отрицательной стороне и его перенос через мембрану на положительную сторону.
Используется при зарядке конденсаторов, базирующихся на цитоплазматической мембране. - Выделение нейтрального атома водорода на отрицательной стороне → его перенос через мембрану на положительную сторону → расщепление на протон и электрон на положительной стороне → обратный перенос электрона через мембрану на отрицательную сторону.
Используется при зарядке конденсаторов, базирующихся как на тилакоидных мембранах, так и на внутренних мембранах митохондрий. - Расщепление на положительной стороне водородосодержащей молекулы у растений и цианобактерий – вода, у пурпурных серобактерий – сероводород с выделением свободных протона и электрона → перенос электрона через мембрану на отрицательную сторону.
Используется только при зарядке конденсаторов, базирующихся как на тилакоидных мембранах.
Во всех трёх способах в результате всех переносов заряд конденсатора увеличивается на одну элементарную единицу е и производится работа
w = еV (4)
по преодолению сил электрического поля при перемещении заряженной частицы через мембрану.
В третьем способе для расщепления водородосодержащей молекулы требуется дополнительная затрата энергии w1 (в пересчёте на один освобождаемый протон), так что полная потребность в энергии для увеличения заряда конденсатора на одну элементарную единицу составляет
w = еV + w1. (5)
Порции энергии, поступающей от внешнего источника и используемой при зарядке конденсатора
Энергия электромагнитного излучения
Дата последнего обновления: 2010-09-09
Файл требует завершения. Предполагается описать более подробно физические процессы, протекающие при зарядке конденсаторов. Далее находятся некоторые заготовки – фрагменты будущего текста
//////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
Перемещение электрона из люмена в строму на этпе 2 производится против сил электрического макрополя и требует затрат энергии
Е l→s = e ∙V, (1)
где е – заряд электрона, V – разность потенциалов стромы и данного конкретного люмена.
* Перемещение электронов из люмена в строму является многошаговым процессом, на каждом шаге которого один электрон «поднимается» от «нижележащей» молекулы КПЭ к «вышележащей».
* Энергия Еliftk, затрачиваемая на k-ой ступени подъёма для преодоления сил электрического макрополя, пропорциональна высоте ступени Δhk = zk+1 – zk для рассматриваемого шага:
Еliftk = Δhk/h ∙Еlift. (2)
Условия насыщения конденсатора
* Как следует из (1) и (4), для успешного переноса заряда необходимо, чтобы энергии используемых экситонов хватило для преодоления сопротивления электрического поля конденсатора:
κ∙ Еq1 ≥ e ∙V для ФС1 (5)
и
κ∙ Еq2 ≥ e ∙V + ½ ЕWS для ФС2. (6)
* Энергия экситонов, не удовлетворяющих условиям (5)-(6), переизлучается в виде люминесценции, в конечном счёте переходит в тепло и бесполезно теряется.
* Из условий (5) и (6) вытекает, что ФС1 перестаёт накачивать энергию в конденсатор, когда напряжение на обкладках превышает величину
V1* = κ∙Еq1/e, (7)
а ФС2 – величину
V2* = (κ∙ Еq2 – ½ ЕWS)/е. (8)
* Величины V1* и V2* представляют собой значения напряжения насыщения для ФС1 и ФС2 соответственно.
* Если справедливо неравенство (3), то
V2* < V1*, (9)
т.е. ФС2 перестаёт перекачивать заряд между обкладками раньше, чем ФС1.
* ФС1 может продолжать перекачивать заряд при неработающей ФС2 только при условии, что задействованы альтернативные пути регенерации ПД1.
* Максимальная величина Е* запасаемой в конденсаторе энергии определяется формулой
Е* = ½ С (V1*)2, (10)
где С – ёмкость конденсатора.
Энергия Е2, затрачиваемая в ФС2 при увеличении заряда конденсатора на одну элементарную единицу, включает в себя, кроме энергии Еlift, необходимой для подъёма электрона с положительной обкладки на отрицательную, также половину энергии ЕWS, необходимой для расщепления молекулы воды:
Е2 = Еlift + ½ ЕWS. (5)
//////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
У цианобактерий тилакоиды образуют более сложные структуры, а роль стромы играет цитоплазма.
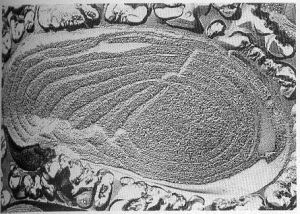
Клетка цианобактерии, внутри которой видны мембраны тилакоидов (из книги LIFE. Science of Biology)
Физический принцип зарядки аккумулятора
В равновесной электрически нейтральной воде имеется одинаковое количество гидроксильных анионов ОН– и катионов гидрония (гидроксония) НзО+.
При зарядке конденсатора ЛСТМ происходит уменьшение содержания гидрония в строме и соответствующее увеличение в люмене.
Процесс зарядки конденсатора может происходить в двух вариантах: зарядка с простым циклом и зарядка со сложным циклом.
Зарядка с простым циклом используется как альтернативный способ в хлоропластах растений и как единственный механизм – в тилакоидах гетероцист (азотофиксирующих клеток нитчатых форм цианобактерий) и фотоэнергетических (фототрофных) бактерий.
Нитчатая цианобактерия Anabaena.
Наиболее многочисленные зеленоватые клетки – вегетативные. Клетка округлой формы слева – гетероциста.
Зарядка со сложным циклом является главным способом для тилакоидов растений и вегетативных клеток цианобактерий.
Зарядка с простым циклом
Принципиальная схема процесса
Установлено, что определяющую роль в структурной реализации зарядки с простым циклом играют два встроенных в тилакоидную мембрану макромолекулярных комплекса: цитохромный комплекс (ЦК) и фотосистема-1 (ФС1).
Нужен рисунок, на котором изображены тилакоидная мембрана, ЦК, ФС1, а также реакции, движение электрона и атома Н на различных этапах.
Простой цикл состоит из шести этапов:
1) электрон, полученный на последнем этапе предыдущего цикла на люменальной стороне ЦК, переносится в люмене от ЦК к ФС1;
2) доставленный к люменальной стороне ФС1 электрон перемещается из люмена в строму по комплексу переноса электронов (КПЭ), являющемуся частью ФС1;
3) перенесенный в строму электрон перемещается по стромальной стороне тилакоидной мембраны от ФС1 к ЦК;
4) доставленный к стромальной строне ЦК электрон е– соединяется с изъятым из стромы гидронием НзО+, что приводит к появлению молекулы воды Н2О и атома водорода Н;
5) атом Н переносится внутри тилакоидной мембраны со стромальной строны ЦК на люменальную.
6) находящийся на люменальной стороне ЦК атом водорода Н расщепляется белками ЦК на электрон е– и протон Н+, который соединяется с молекулой воды из люмена, образуя гидроний НзО+;
Далее происходит переход к первому этапу следующего цикла.
Гипотеза о подвижных молекулах-переносчиках
Согласно общепринятой теории этапы 1 и 3 простого цикла реализуются подвижными белками-переносчиками электрона, непрерывно курсирующими в процессе пассивной диффузии между двумя пунктами – посадочными местами на сответствующей строне ЦК и ФС1.
Роль белков-переносчиков в этой теории играют:
- на этапе 1 – катионы пластоцианина Cu+Pc (для цианобактерий – цитохрома Fe2+с6);
- на этапе 3 – анионы ферредоксина Fd–.
// Молекулы пластоцианина и ферродоксина содержат порядка 1000 атомов и имеют молекулярную массу порядка 104 Д.
// Молекулы пластоцианина имеют размеры, сопоставимые с толщиной слоя люмена, в котором они должны диффундировать.
По некоторым предположениям перенос атома водорода на этапе 5 осуществляют подвижные молекулы пластохинона РQ, перемещающиеся внутри тилакоидной мембраны от стромальной ЦК к люменальной стороне и обратно в ходе пассивной диффузии;
// молекулы пластохинона содержат 128 атомов и имеют молекулярную массу 675 Д.
Гипотеза о подвижных макромолекулах-переносчиках нефизична и должна быть отвергнута.
Гипотеза, представляющаяся более реалистичной
Более реалистичным представляется предположение о том, что посадочные места на ЦК и ФС1 со стороны люмена соединены выстроенными в цепочку неподвижными молекулами пластоцианина, а со стороны стромы – такой же цепочкой молекул ферредоксина.
Передача электрона может при этом происходить от одной молекулы к другой вместе с передачей энергетического возбуждения в виде солитона.
Перемещение электрона происходит при этом в областях, где макрополе отсутствует, а микрополе имеет периодический характер. Таким образом, на каждом шаге передачи затраты энергии в микрополе чередуются с такими же по абсолютной величине её приобретениями при общем нулевом балансе.
Аналогично, на этапе 5 перенос нейтрального атома водорода могут осуществлять выстроенные в цепочку молекулы пластохинона.
Баланс одного оборота простого цикла
- движение электронов происходит по замкнутому контуру, так что их количество в люмене и в строме остаётся неизменным.
- в строме количество протонов уменьшается на единицу, а отрицательный заряд увеличивается на элементарную единицу;
- в люмене количество протонов и положительный заряд увеличиваются на единицу.
Зарядка конденсатора производится путём переноса протонов из стромы в люмен.
Потребности в энергии для процесса зарядки с простым циклом
Поскольку каждый протон переносится из стромы в люмен на этапе 5 совместно с электроном в составе нейтрального атома водорода, этот перенос не требует совершения работы против сил макрополя конденсатора.
Перемещение электрона из люмена в строму на этпе 2 производится против сил электрического макрополя и требует затрат энергии
Е l→s = e ∙V, (1)
где е – заряд электрона, V – разность потенциалов стромы и данного конкретного люмена.
На этапе 4 в реакции соединения гидрония с электроном выделяется энергия, равная работе их микрополей при сближении протона и электрона.
На этапе 6 при расщеплении атома Н требуются затраты энергии на разведение электрона и протона (энергия ионизации).
- На каждом энергопотребляющем этапе цикла должна подводиться необходимая энергия к устройству, реализующему этот этап.
- В частности, должен быть обеспечен непрерывный подвод энергии к каждому элементу КПЭ и к расщепляющему атом водорода устройству на люменальной стороне ЦК.
└──┘ └──┘ //
ВРК ФНР //
// └───────┘ └─────┘ └──────┘ └───────┘
// ФС2 ЦК ФС1 АТФ-аза
Схема тилакоидной мембраны и встроенных в неё устройств
(Рисунок взят на сайте: Robert Carpentier: http://www.uqtr.ca/labcarpentier/eng/home_frames.htm )
Пошаговое распределение затрат энергии
* Перемещение электронов из люмена в строму является многошаговым процессом, на каждом шаге которого один электрон «поднимается» от «нижележащей» молекулы КПЭ к «вышележащей».
* Энергия Еliftk, затрачиваемая на k-ой ступени подъёма для преодоления сил электрического макрополя, пропорциональна высоте ступени Δhk = zk+1 – zk для рассматриваемого шага:
Еliftk = Δhk/h ∙Еlift. (2)
* Одна часть пути, который проходит электрон при подъёме от люмена к строме, лежит ниже ФРЦ, а другая часть – выше.
Баланс энергии для элементов КПЭ
* Каждому вновь поступающему к ФРЦ солитону, обладающему достаточной энергией, соответствует свой цикл передач электронов от люмена к строме.
* В случае установившегося режима работы для каждого элемента КПЭ:
- потери энергии Еdetk при отделении электрона компенсируются выигрышем энергии за счёт работы микрополя при присоедении электрона во время предыдущего цикла передач.
- работа Еliftk, затраченная на подъём электрона в макрополе, должна быть компенсирована энергией Еsolk, принесенной солитоном.
- с учётом неизбежных потерь энергии Еlossk баланс энергии требует выполнения равенства
Еsolk = Еliftk+ Еlossk. (3)
* В момент начала зарядки аккумулятора на восходе Солнца:
- ввиду отсутствия предыдущего цикла передач потери энергии при отделении электрона Еdetk должны компенсироваться энергией Еsolk, принесенной солитоном;
- макрополе и связанная с ним работа Еliftk равны нулю;
- баланс энергии имеет вид
Еsolk = Еdetk+ Еlossk. (4)
* Описанный порядок передачи энергии и величина передаваемой части должны быть предопределены структурой передающей и принимающей энергию молекул.
// Направление потока энергии не всегда совпадает с направлением потока электронов.
// В рассматриваемом процессе электроны не являются переносчиками энергии.
// Термин высокий энергетический уровень относится к полной энергии электрона, включающей как потенциальную, так и кинетическую энергию, и, следовательно, имеет смысл только для электрона в составе атома.
// Покидающий атом электрон уносит энергию, состоящую из двух слагаемых, не зависящих от предыдущего энергетического состояния:
- энергия создаваемого электроном магнитного поля, имеющая фиксированную величину;
- собственная кинетическая энергия электрона, как правило, весьма небольшая.
// Распространённое представление о «высокоэнергетичном» электроне, передающем по частям свою энергию при переходе от одной молекулы к другой, физически некорректно.
* В случае успешного завершения цепочки пошаговых подъёмов электрона люмен теряет, а строма приобретает один элементарный отрицательный заряд.
* На осуществление процесса переноса одного элементарного заряда тратится энергия Еsol солитона, порождённого одним квантом солнечного излучения.
Энергия Е2, затрачиваемая в ФС2 при увеличении заряда конденсатора на одну элементарную единицу, включает в себя, кроме энергии Еlift, необходимой для подъёма электрона с положительной обкладки на отрицательную, также половину энергии ЕWS, необходимой для расщепления молекулы воды:
Е2 = Еlift + ½ ЕWS. (5)
Общепринятые модели регенерации конечных акцепторов и первичного донора ФС1
* Предполагается существование подвижных молекул-переносчиков электрона, непрерывно курсирующих в процессе пассивной диффузии между двумя пунктами – посадочными местами на своей фотосистеме и на соответствующем устройстве для регенерации.
* В случае ФС1 роль переносчиков электрона должны играть:
- для первичного донора – белковые катионы пластоцианина Cu+Pc (для цианобактерий – цитохрома Fe2+с6), курсирующие в люмене между нижней стороной ФС1 (место отдачи электрона) и нижней стороной цитохромного комплекса (ЦК), где электрон приобретается;
- для конечного акцептора – белковые анионы ферредоксина Fd–, которые курсируют в строме между верхней стороной ФС1 и ферредоксин-NADP-редуктазой (ФНР) (альтернативный путь – между верхними сторонами ФС1 и ЦК).
* Молекулы пластоцианина и ферродоксина содержат порядка 1000 атомов и имеют молекулярную массу порядка 104.
* Играющие роль конечного акцептора для ФС2 молекулы пластохинона РQ.
внутри плотно заполненной белками тилакоидной мембраны между ФС2 и ЦК
* Молекула пластохинона имеет 128 атомов и молекулярную массу 675 Д.
* При регенерации КА2 – пластохинона РQ – требуется изъятие двух электронов у возникшего в конце рабочего цикла аниона РQ2–, что достаточно проблематично в отрицательно заряженной среде стромы.
* Проблемы регенерации решают в рассматриваемом случае встроенные в мембрану тилакоида цитохромные комплексы b6/f (ЦК).
* Согласно общепринятой в настоящее модели процесс регенерации КА2 состоит из следующих этапов.
- Образовавшийся после приёма двух электронов анион РQ2– отделяется от ФС2 и перемещается вдоль поверхности мембраны тилакоида к первому посадочному месту на ЦК, расположенному со стороны стромы.
- На первом посадочном месте РQ2– соединяется с двумя протонами Н+, полученными из пула стромы, передавая тем самым строме двойной отрицательный заряд, и превращается в электрически нейтральную молекулу пластохинола РQН2.
// В другом варианте модели присоединение протонов и превращение в пластохинол происходит до отделения пластохинона от ФС2.
//Гипотеза о наличии в отрицательно заряженной строме двух свободных протонов, причём в нужное время и в нужном месте, не кажется очень убедительной. Более вероятным представляется извлечение из стромы нейтральной молекулы Н2, её разделение на два протона и два электрона, присоединение протонов к пластохинону и передача электронов в пул стромы.
- Молекула РQН2 отделяется от первого посадочного места на ЦК и перемещается на второе посадочное место, расположенное непосредственно у поверхности люмена.
// Перемещение нейтральной молекулы от отрицательной обкладки конденсатора к положительной не требует затраты энергии. - Новое посадочное место является исходным положением для 4-х стадийного процесса отбора атомов водорода от молекулы РQН2.
- На каждой стадии обрабатываемая молекула перемещается от одной белковой субъединицы ЦК к другой, последовательно оставляя на них два электрона и два протона и превращаясь в итоге в исходную молекулу пластохинона РQ.
- Нейтральная молекула РQ отделяется от ЦК, перемещается внутри мембраны тилакоида или по поверхности мембраны со стороны люмена к ФС2, поднимается по внутримембранному пути к своему посадочному месту на ФС2 и садится на него.
- Два свободных протона переходят в пул люмена, а электроны остаются связанными на белковых молекулах ЦК.
* Предполагается, что все перемещения молекул пластохинона и пластохинола происходят в результате пассивной диффузии.
Цикл регенерации ПД1
* Первичными донорами для ФС1 являются катионы ПД1+, имеющие один электрон в валентной оболочке атома меди или железа.
* Для растений роль ПД1+ играет катион пластоцианина Cu+Pc, а для цианобактерий – катион цитохрома Fe2+с6.
* В процессе работы ФС1 молекулы ПД1+, отдавая последний валентный электрон атома меди или железа, увеличивают свой положительный заряд и превращаются в катионы ПД1++ (соответственно Cu2+Pc и Fe3+с6).
* Регенерирующими устройствами для ПД1+ являются те же ЦК, что и для КА2.
* После срабатывания ФС2 катион с увеличенным зарядом ПД1++ отделяется от ФС1 и перемещается в люмене вдоль поверхности мембраны на своё посадочное место в ЦК.
* В общепринятой модели:
если ФС2 срабатывает чаще, чем ФС1,
(временные промежутки t1 и t2 удовлетворяют условию t1 ≥ t2),
то
////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
ЦК передаёт ПД1++ один из электронов, оставшихся у него после обработки КА2, возвращая его в состояние ПД1+.
//* Если какой-либо электрон из оставшихся на ЦК после регенерации КА2 оказывается невостребованным до прихода следующего электрона, он переходит с ЦК в пул люмена, соединяется с протоном из пула и превращает его в нейтральный атом водорода.
/////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
в противном случае, т.е. когда ФС1 срабатывает чаще, чем ФС2,
/////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
Работает альтернативный механизм регенерации ПД1+, состоящий в том, что ЦК передаёт ПД1++ //электрон,
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
* Регенерированный катион ПД1+ возвращается по обратному пути в люмене к своему посадочному месту на ФС1.
Регенерация КА1
* КА1 является молекула ферредоксина Fd, обращающаяся после приёма электрона в анион Fd–.
* Регенерация КА1 сводится к нейтрализации аниона Fd–, т.е. к изъятию у него лишнего электрона.
* Отдача электрона в отталкивающую его отрицательно заряженную среду требует использования специального оборудования, которое предоставляет встроенная в мембрану тилакоида по соседству с ФС1 и обращённая к строме ферредоксин-NADP-редуктаза (ФНР).
* В настоящее время существуют две конкурирующих модели процесса регенерации ферредоксина:
- анион Fd– переносится в строме к ФНР, где нейтрализуется, а затем возвращается на своё посадочное местов ФС1;
- находясь на ФС1, ферредоксин расположен так близко к ФНР, что процесс регенерации происходит без всяких перемещений.
/////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
Условия насыщения конденсатора
* Как следует из (1) и (4), для успешного переноса заряда необходимо, чтобы энергии используемых экситонов хватило для преодоления сопротивления электрического поля конденсатора:
κ∙ Еq1 ≥ e ∙V для ФС1 (5)
и
κ∙ Еq2 ≥ e ∙V + ½ ЕWS для ФС2. (6)
* Энергия экситонов, не удовлетворяющих условиям (5)-(6), переизлучается в виде люминесценции, в конечном счёте переходит в тепло и бесполезно теряется.
* Из условий (5) и (6) вытекает, что ФС1 перестаёт накачивать энергию в конденсатор, когда напряжение на обкладках превышает величину
V1* = κ∙Еq1/e, (7)
а ФС2 – величину
V2* = (κ∙ Еq2 – ½ ЕWS)/е. (8)
* Величины V1* и V2* представляют собой значения напряжения насыщения для ФС1 и ФС2 соответственно.
* Если справедливо неравенство (3), то
V2* < V1*, (9)
т.е. ФС2 перестаёт перекачивать заряд между обкладками раньше, чем ФС1.
* ФС1 может продолжать перекачивать заряд при неработающей ФС2 только при условии, что задействованы альтернативные пути регенерации ПД1.
* Максимальная величина Е* запасаемой в конденсаторе энергии определяется формулой
Е* = ½ С (V1*)2, (10)
где С – ёмкость конденсатора.
* Группировка тилакоидов в стопки имеет тот же смысл, что и в случае обычной батареи конденсаторов – появляется возможность при том же напряжении насыщения получить бóльшую запасённую энергию.